Deep Shade and Velvety Leaves
This post is unusual, and certainly not of universal interest! But it represents what I’ve been chewing on for a few days, so what the hay, it relates to native plants in a broad way.
Our local Dull Leaf Coffee doesn’t have dull leaves it all. They have a lustrous velvety sheen, and are an example of a widespread adaptation of many understory shade species. Gardeners may be familiar with similar leaves on begonias, on aroids, and on shooting star clerodendrum.

Dull Leaf Coffee with velvet leaves. (By John Bradford)
Those velvety surfaces reveal a microscopic array of lenses and solar collectors so “optical” that such leaf surfaces have been studied from the standpoint of lenses. Florida can claim the modern world’s authority on leaf optics, although 19th Century German botanists had a good go at it. Today’s material springs from a couple publications by FIU Prof. David Lee with an assist from “The Life of a Leaf” by Duke University Prof. Steven Vogel. As Dr. Lee notes, it’s not all figured out! But there’s plenty to discuss, and don’t blame on them shortcomings in my interpretation.
How often do we stop and think of the diversity of leaf surfaces? What’s more conspicuous in the plant world than all those shades of green, variegations, textures, sheens, twinkles, and lusters? That green rainbow has meaning.
Some deep-shade species have a blue iridescent aspect, uncommon in Florida native plants, although visible in garden species (for instance Selaginella uncinata shown below) and sometimes in young native Virginia Creeper. Such iridescent species absorb useful light especially effectively while reflecting wavelengths we perceive as near-blue.

Leaves apparently use “fiber optics” along their cell walls to transmit light from the surface down deep. Purplish layers toward the bottom of the blade seem to backscatter escaping pass-through light back up into the photosynthetic region for a double dip. Good examples include some begonias, prayer-plants, zebrina, oyster plant, and shooting star clerodendrum.
To get now to today’s feature, velvety shade leaves have convex lens cells on top. They focus incoming light so sharply that bright focal points are sometimes visible through a microscope from these little magnifying glasses. Additionally, the cells refract into the leaf light arriving at diffuse angles. The drawing below from a paper by Dr. Lee from the 1980s shows a lens cell with rays arriving at an angle, refracted, and focused into the cell and on downward. Light arriving obliquely might otherwise be more wasted reflecting off the surface.
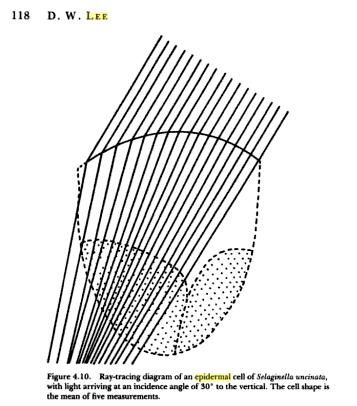
The photo below shows lens cells on the surface of a Dull Leaf Coffee.

You might ask, if a certain amount of light enters a leaf, why focus the dose into a smaller brighter beam? And to sharpen that question: Sometimes even under the dense canopy light can be too bright, say during fleeting moments of sun flecks, or maybe dawn or dusk as the sun peeks in. Couldn’t bright focused beams then be excessive? Probably, and one protection might be variegation, which we’ll consider in a moment.
There’s more to light entry than merely striking a static surface. The leaf adjusts internally to light intensity. The sun beams strike tiny Frisbees, chloroplasts, within the layered leaf cells. Chloroplasts in modest light cluster on the horizontal floor of the cells with their broad faces exposed to maximize capture. Think of closed venetian blinds. When the light is too strong, however, the chloroplasts migrate away from the light and align themselves along the vertical cell walls facing the illumination with their narrow edges. Open venetian blinds. When the blinds are open a bright focused beam reaches into deeper layers. Thus a benefit of a focused beam might be controlled penetration into the blade with overall great effect.

In the illustration above, the two leaf cells on the left under a lens cell have their chloroplasts oriented face-up for maximum light capture. On the right, the more intense focused beam penetrates as the chloroplasts orient safely edge-wise, capturing the light eventually, just deeper into the leaf.
It is possible demonstrate chloroplast adjustment by covering part of a shade leaf, then shining light on it. The covered portion stays dark green with its chloroplasts in their “shade orientation,” while the exposed portion lightens in color as the chloroplasts adjust into the “open blinds” orientation.
Leaf Variegation
Gardeners oh so love variegated leaves. Although I think they are pretty, they don’t send me into raptures, maybe due to my preference for native species in landscaping, with so few natives being variegated. To my eye a highly variegated garden tends to look contrived and unnatural. We do have a few though: some Smilax, for instance.

Smilax dabbed with the brush. (By John Bradford)
There’s no single known answer for, “what’s variegation good for,” and little hard data. I’m less confident of the discussion that follows than of the one above. Brace for rampant speculation.
Variegation, defined as patterns in leaf coloration, has arisen many times in many ways. Some of it comes from viral infections, probably no benefit to the plant. This is the case for the so-called Flowering-Maple, Abutilon pictum. Many other variegated plants owe their patterns, usually streaks or blotches, to minor mutations during foliar development. Again, if these are beneficial it is not clear how or why.
But some variegation is clearly genetic, heritable, complex, patterned, and almost certainly usefully adaptive. But even that is tough to interpret. Botanists have considered various explanations, which are not mutually exclusive, and which may apply differently, combined, or not at all to different species. Ideas about leaf coloration patterns include signals to herbivores of poor nutritional quality or of toxicity (so leave me alone); or false damage making it look to herbivores like a different pest beat them to the salad bar; or as camouflage to throw off the herbivore’s innate “search image.’” Some observers have suggested leafy colorifics to attract pollinators.
Such critter-based hypotheses may hold water, remembering especially that variegation has arisen many times , but the creature-centric interpretations don’t take into account an apparent correlation between variegation and dark shaded habitats.
Variegation in deep shade presents an apparent dilemma: variegation diminishes the photosynthetic surface, seemingly not a “good idea” for the very shade-bound plants prone to patterned leaves. But here’s a possible explanation, neither original with me nor “accepted fact”: deep-shade leaves are good at catching light when the intensity is low, but intensities above the usual gloom can impair photosynthesis, especially in leaves “designed” for low light, including those with lens cells. Every gardener knows that shade species popped into bright sun suffer.
Those shade-adapted leaves are more delicate than sun-loving leaves, dandy down where it is dark, moist, and still. But intermittent bright light may be a problem as winds blow, as the shifting sun peeks under the canopy, as sun flecks dance through the foliage. Or where light exposure changes with plant age and stature.
Variegation may sometimes give shade-leaves protection from sporadic sunny moments.
Some plants, such as some Cecropias, tend toward variegation when young and shaded in the understory but not variegated when older, taller and sunnier. As David Lee illustrated, at least one African-Violet has its variegation increase as the day progresses.
It may be—and there is need for far more research—that some leaves with a varied coloration pattern enjoy the best of both worlds, the non-variegated portions function best in low light, with variegated portions less light-sensitive and thus happier at the brighter moments. A leaf able to function well only in deep shade may waste a lot of photosynthetic opportunity at brighter times, but a two-toned job is ready for anything.
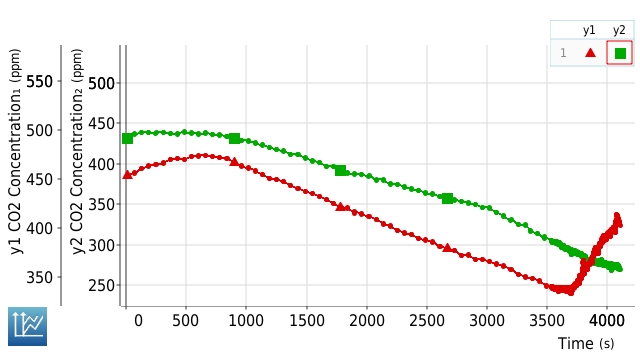
Image above: Apparent advantage of variegation. This graph shows two tissue regions from the same Marnata (Prayer Plant) leaf monitored simultaneously under changing laboratory light exposure conditions (adjustable lamps). The vertical axis shows photosynthesis/respiration as CO2 usage/release. The horizontal axis shows seconds with increasing light intensity. (And heat increased with light intensity.) The top line (Y2) shows performance by the variegated portion of the leaf. The bottom line (Y1) shows the unvariegated green portion. Under low light they behave similarly, with the green unvariegated portion slightly more responsive to the increasing light. At about 3600 seconds the increasing light and associated warmth caused the unvariegated portion to stop constructive photosynthesis and switch to destructive respiration, releasing carbon dioxide abruptly. The variegated tissue passed that threshold unscathed, clearly standing up more successfully to the increased simulated sun exposure
John's and my blog this week








































{kind=link}