Note added in 2020: The original version of this post dates back a few years. It has been updated repeatedly due to new thoughts and new information. If you are looking for a definitive proven answer on the adaptive significance of cypress knees, not here. Or not yet.
If you’d like a short answer, personally I strongly suspect the knees allow the living tissues of the root to come up for air, providing a large surface area for the vascular cambium and more importantly the sugar-pumping (and possibly storing during the leafless season) phloem, like surface pumping stations along a pipeline. That is by far my preferred hypothesis, and I’m trying every low-tech way to test it I can. But much fun stuff to discuss, read on…
Observations, speculations, interpretations and questions about cypress knees date back about as far as botanical writing concerning the Southeastern U.S. And beyond:
Any interpretation of the knees must extend beyond Taxodium distichum in and near the Southern U.S. Other Taxodiaceae/Cupressaceae, including Asian representatives, can form knees. Whatever functions the knees serve must pertain broadly in time, space, and related species.
It is surprising, after centuries of interest, how much interpretation is based on field observation, and how little hard data exist on knee anatomy, cellular structure, and physiology. Not easy to study, let alone to monitor the growth of woody glaciers. As field equipment becomes smaller, less expensive, and accessible, we may see a new development of data to help settle questions. The phloem physiology of Taxodium roots and knees has a target on it.
And while in the realm of ifs, and, and buts, it may be worthwhile to mention that multiple functions are possible, even if we prefer one adaptation as “the” main driving force behind knee evolution. One function does not necessarily preclude others. And also conceivable is no particular function.
A wonderful summary of what came before and an in-depth new analysis is by H. S. Lanborn in the American Naturalist 1890. There seems to have been a flurry of interest in late 19th Century botany. Interestingly, after airing the evidence thoroughly, including the common observation that knees tend to form where aeration is needed, Lanborn disfavored the usual interpretation of cypress knees as air-exchangers in preference for a pair of alternative hypotheses….that the knees function primarily to brace the trees against root failure during storms and to catch floating debris, which would add weight on top of the root system.
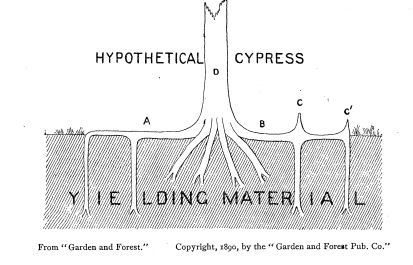
Lanborn drew the diagram above, showing how he felt the knees with their broad bases reinforced the roots (root B stronger than root A). Lanborn recognized that the knees rise far higher than needed to serve as root reinforcements, which led him to the secondary hypothesis of the pegs as flotsam-catchers. The brace-flotsam interpretation thus has a fundamental problem by invoking two functions.
Long before and after Lanborn, the conventional interpretation of the knees is as ventilation devices. Cypress roots run horizontally through anoxic mud and water and thus need air. Other wetland plants have aerenchyma tissue allowing gas exchange between deep roots and the soil surface (although problematically such tissue is unknown in Bald Cypress). Despite tilting away from ventilation, Lanborn and others, including me, agree that knee formation seems to correlate (here’s a chance for quantified research) with growth where roots would otherwise be smothered under water or anoxic mud. Assuming this correlation stands up to systematic study, it is in itself evidence for a role in gas exchange somehow. After all, the knees are called “pneumatophores.”
As Lanborn quoted an earlier observer:
“It seems likely, therefore, that some process connected with the exposure of the sap to the air takes place in these protuberances.”

Solid wood. (I did not cut the knee.)
The ventilation concept has complications. At first glance, it is easy to suspect the knees are the above-water snorkels to a generalized root aeration system. The knees are wood with no apparent ductwork. The water-conducting cells are tracheids, which have a membrane (pair of adjacent cellulose cell walls) blocking the passageways (pits) from cell to cell. The pit membranes are not air-tight but there are millions of them in the path of the gas exchange. But are the knees completely air-tight? Not necessarily.
First of all, aging knees can become hollow with age, obviously then allowing air to pass readily to the base of an older decaying knee. Further, a recent study shows increased oxygen in Taxodium roots near the bases of exposed knees as opposed to under submerged knees. CLICK for an abstract. Gases do diffuse in wood or be dissolved in water or phloem sap, but very slowly, and remember, the knees are a long distance from the young growing feeder roots, and the water movement in the wood, at least when the tree is active with foliage present, is in the wrong direction.
This study you just clicked contradicts earlier research where aeration was not in evidence. The recent positive results are based on needle-drawn air samples from root wood near the knee bases. The experiments dug and cut free knees with portions of root attached and transported the knees to a lab, then studied the knees in an aquarium many hours later under varied water depths. The experimental technique complicates interpretation of the results:
Most importantly, root xylem under natural conditions usually has negative pressure, “suction,” as transpiration pulls water and any dissolved oxygen inward to the trunk and up the tree, away from the feeder roots. The instant the roots are severed the negative pressure is eliminated, and any related gas dynamics are lost, making it tough to know what to make of oxygen levels in such depressurized dying roots.
Further, it is not clear if the needle-drawn air samples were consistent in origin, and each from sapwood, heartwood, wood rays, or pith. Root damage and changing conditions could elicit a hormonal response with metabolic consequences. It was not clear if the knees used were solid, partly decayed, or even hollow (probably not, but their structural condition matters). It was also unclear if oxygen entry was via wood, or alternatively along the porous bark we’ll return to momentarily. Maybe even from air trapped in the bark during transport to the lab.

By John Bradford
Despite all these worries, my hunch is to agree that some oxygen may well reach the base of the knee and attached adjacent root. But then a bigger question: exactly what living tissue (wood is mostly non-living), and where does the oxygen serve? Perhaps oxygen makes its way along the length of that woody root snaking through the mud in ways not adequately explained anatomically, and then somehow ventilates the system. The concept of effective long-distance air movement in the wood is problematic.
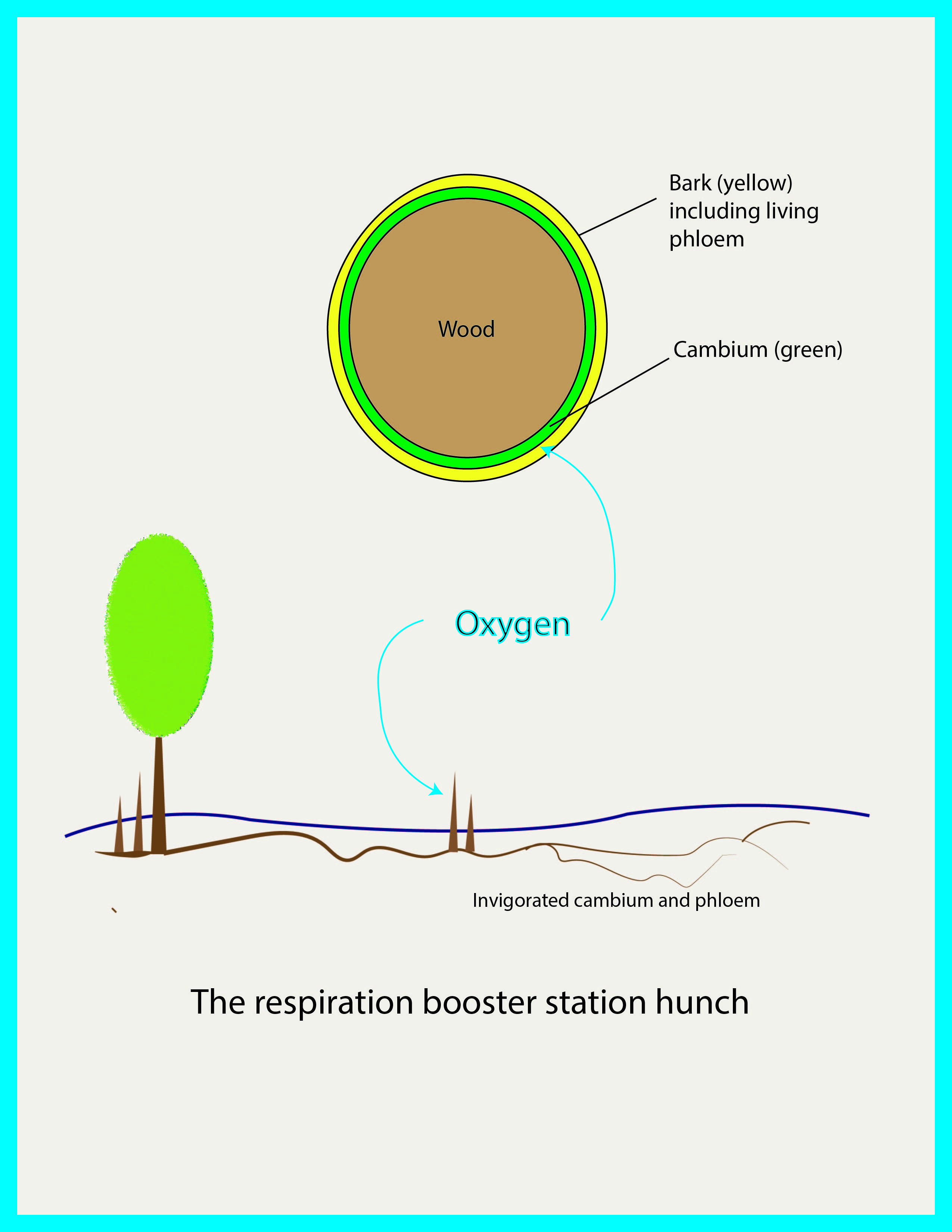
But something else moves the length of the root through known channels under pressure and requires oxygenation: the phloem sap. So maybe the oxygen entering the knee (and possibly reaching adjacent roots) is most important at and near the knee, not to ventilate the root, but rather as a periodic surface-oxygenating rejuvenation station for the phloem, a pumping station.
There is a vague and inchoate hint of this as far back as Lanborn. Review the quote dating to 1879 above. It does not refer to aerating the root, but rather to exposing the sap to the air. The root system pumps sugary phloem sap all the way from the leaves out to the tips of the tiny feeder roots. This requires oxygen. Oxygen is in short supply under that water and wet mud. Knees rising periodically and exposing the phloem (and associated cambium) at the knee surface to a periodic breath of fresh air may keep the sap flowing.

Knees often form on arched roots.

A photo my dent1st would love (exposed roots). This big old knee, and many other like it, does not say “snorkel” to me. It says “root hub.” It sure could be a place where radiating roots get a dose of life-giving gas exchange.
The phloem is a continuous living sleeve covering the entire root. It has two layers: a soft spongy outer dead layer (bark) covering the living breathing pressurized inner layer where the sugary sap flows. Just under that is the likewise living vascular cambium responsible for growth in diameter, and all this exterior to the wood. Think of the wood as a wire and the phloem/cambium as the plastic insulation covering the copper.
Air entering the knee would contact and presumably benefit the phloem immediately. The knees has a large surface area potentially exposing a lot of phloem and cambium to a lot of air, a giant gill, with no need to postulate gas exchange through dead wood under negative pressure in the wrong direction.
This would explain why the knees rise so high…to provide a big exposed surface. This fits with the acknowledged prevalence of knees in situations where oxygenation is needed. And localized gas exchange at and near the knee is consistent with the recent observations of oxygen getting into the knees and adjacent roots, diffusing through wood and/or through the porous bark, and maybe even carried along as dissolved oxygen in the reinvigorated outbound phloem sap.


Knees on shore
To continue with the phloem, the trees are leafy much of the year, corresponding more or less to the wet season, and “bald” leafless during more or less the winter months and dry season, although the wet-leafy/dry-bakd correspondence is not exact. In any case, the tree goes months with no photosynthesis. No sugars being made. What does it do about that? In line with the idea that the knees are basically sugar movers, maybe they are in the moving and storage business, serving as sugar reserves, sort of like our livers, in between the intermittently sugar-producing leaves and the always-sugar-needing roots. I am trying to find a way to detect sucrose storage in the knees. A bloof glucoe meter may come in handy although it tests for glucos not sucrose, And of coruse, you do not go to wild areas and start cutting of knees! In the mod 80s biologist Clair Brown used an iodine test to check for starch in cut-off knees, and they were positive. Starch is stored sugar, so you may say her results were not far apart from the present ypothesis. Funny how observations converge.
Even so, I plan to see this idea through ethically and effectively.
There have been additional knee proposals. Here are some:
- Serve as variable-height launchpads for tufts of feeder roots. The idea here is that the water levels rise and fall during the very long life of a bald cypress. As the water level fluctuates small feeder roots might branch off the trunks and knees to exploit the oxygen-rich water at the water surface. I have not seen such tufts personally, although supportive observations are reported. I wonder why so many trees would make so many knees not apparently engaged in this way, seemingly an expensive investment in a rare contingency. I wonder similarly if the cluster of knees could make enough of a contribution to justify the substantial investment in those big woody knees. If the knee does sprout a tuft of feeder roots in the water allowing nutritional supplementation (from water?) this input is separated physically from the exponentially greater mass of feeder roots at the distal end of the root system. The water and dissolved nutrient input from knee-roots would go into the xylem and would be pulled to the trunk by the inward/upward direction of the transpirational xylem stream, feeding the above-ground tree, not the distal root system. Readers interested in exploring this intriguing hypothesis should consult, Stahle, D. et al. Quat. Sci. Rev. 34: 7. 2012.
- Allow methane to escape. (Here is some info on methane release)
- Rise into stumps and extract nutrients (No evidence and no apparent adaptation to this purpose, and knees generally encountered rising into thin air.)
- Food storage (no traction)
- Will grow into new trees (no)
- No function, merely tumors or burls. (See below)
- Giant thorns. (See below)

Cypress knees are often not single units, but rather commonly clustered, hinting that whatever “set off” a knee kicked off multiple knees in one fell swoop. Some are branched, or are the fusion of adjacent knees creating a branched appearance. Often the knees start from the high point of looping roots, as if a giant knee formed on top of the St. Louis Arch (photo above).

The could be burls, reaction wood, hormonal aberrations, or the results of injury. Interestingly, Bald Cypress trunks will grow a burl at points of abrasive contact. The knees are so widespread and so abundant, their existence as mere burls or growth aberrations is not an appealing hypothesis.

Don’t trip over those “knees.” This painting is by Brian Regal (in The Evolution and Extinctions of the Dinosaurs by D. Eastovsky and D. Weishamel. 1996) . This is based on the Triassic Chinle Formation in Arizona, probably earlier than the appearance of Taxodium, although maybe not before its similar ancestors.)
An intriguing if far-fetched possibility is maybe the knees block large herbivores. (They sure block canoes.) Even if unlikely, fun to envision. Members of the Taxodiaceae (or Cupressaceae) date back to dinosaurs, some of them hunky hungry herbivores. More recently and locally, Mastodons ate Bald Cypress in massive quantities. In the Aucilla River in the Florida Panhandle remain to this day literally truckloads, tons, of preserved Mastodon dung. Guess what it is made of mostly: Bald Cypress twigs.

Yep—that’s it. 12,000 years old yet still fresh. (Photo from UF Museum of Natural History)

Notes:
Newsom, L.A., and M. C. Mihlbachler. Mastodons (Mammut americanum) Diet Foraging Patterns Based on Analysis of Dung Deposits. Springer. 2006.
John's and my blog this week














































{kind=link}