Utricularia subulata (U. cleistogama)
Lentibulariaceae
John and George walked in Savannas State Park in Jensen Beach, Florida yesterday. Birding was the primary agenda. The botanical treat was the wetland areas all abloom with yellow Ludwigias, Sneezeweeds (Helenium—wow, like a garden), Tickseeds (Coreopsis), Xyris, and carnivorous Bladderworts. Some time ago in this blog we looked at the bladderwort trap so we’ll jump past that now.
The Zigzag Bladderwort, presumably named for its zigzag stolon, is kinky in several additional ways:
First, it lives, not generally in standing water like the other local Bladderworts, but rather in moist soil, and trends a little weedy. Look in disturbed moist depressions along wet foot trails.

Zigzag Bladderwort as it looked yesterday. (First three photos by John Bradford.)
Second odd feature, Zigzag BW has by far the broadest distribution of any U.S. Utricularia, extending to Nova Scotia, to Lake Michigan, and to Tennessee. You might ask, how does this delicate little wisp survive cruel ice and snow? It creates subterranean “buds” able to survive tough times.

ZZBW resembles a Snapdragon
Odd item number three, “sticky dew drops” sometimes at branch points. Why would it do that? Maybe to prevent ground-dwelling creepy-crawlies from messing with the blossom? The droplets are super sticky stretchy, natural flypaper.

Note the sticky droplet. These occur sometimes on the stems, where tiny insects (?) can be seen to crawl. (Photo by John Bradford)
If you touch one and pull your finger away a strand stretch to connect your finger to the stem, like gum on a shoe. The droplets could be extrafloral nectaries, ant feeders. However, the extreme tackiness makes me vote for bug-catcher. A second level of blossom security is apparent. Like a Snapdragon, the flower is closed at the mouth until a bee forces the two lips apart.

Look closely—cooties!
Peculiarity four, along the stem, including the subterranean stolon, there appear tiny polyps. I don’t know what these are or why they exist. Other species of Utricularia have gland-tipped hairs, and the bumps on U. subulata look like eensie weensie plant hairs (marked with red circles in the photos). Maybe they are glandular.

The entire plant. It’s all flower!
Weird item five, there are not roots. The underground portion is stem (stolon), and there’s precious little of that. It has a string of nearly microscopic bladder traps. The aboveground stems are mere threads usually under 6 inches tall. The leaves are puny and inconspicuous. In short, it is somewhere between awesome and mysterious that a rootless and nearly leafless plant can rustle up enough “fertilizer” nutrients and sufficient photosynthetic nutrition (carbohydrates) to crank out big booming blossoms. There are factors in this equation we don’t yet know. So read on to fill the gaps with speculation.

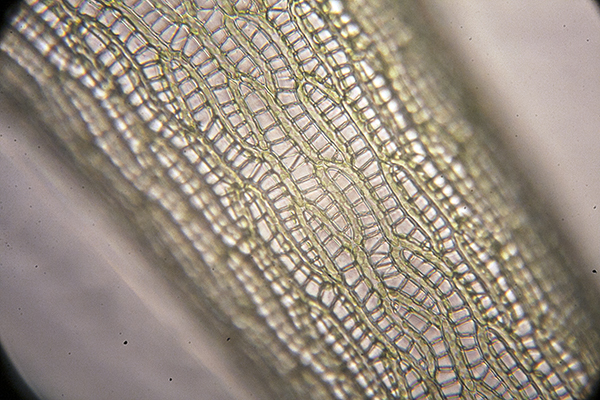
Stolon highly magnified, with traps.
Carnivory in plants is generally presumed to supply “fertilizer” nutrients, mostly nitrogen. The Bladderwort traps do at least this. Utricularia traps come with glands to remove excess water from the closed chamber, and different glands to secrete enzymes into the trap and absorb the digested prey back out, like my stomach. That’s all old-school.

“Hair” on stem. There is another at the bottom of the picture. I don’t know what these are about. Similar structures are on the stolon.
New research hints that the traps are complex little microbial ecosystems perhaps making surprise contributions beyond mere nitrogen. Mainly, carnivory sometimes probably captures and contributes carbohydrate energy to the plant, that is, helps out surprisingly in the “photosynthesis” department beyond the expected “fertilizer” department. Some Utricularia species actively feed symbiotic trap-dwelling microbes, which may help lure prey, digest the victims, and (here’s the stretch) somehow transfer carbohydrate energy stolen from the prey to the plant. If true, that would help explain how the plant drums up enough carbohydrate-ish energy despite being underendowed photosynthetically.
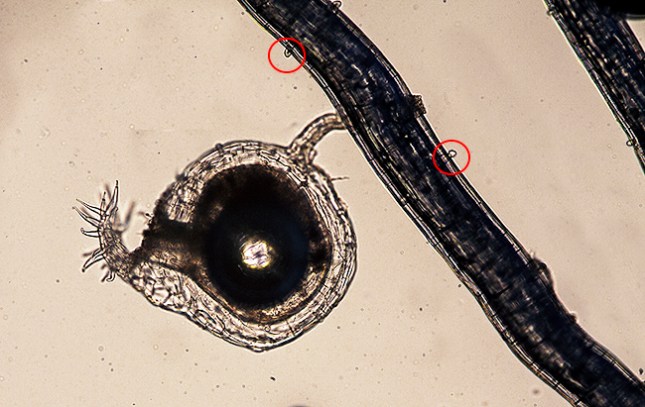
The trap under a microscope. The trap entrance (left side) and its “welcome pricker thing” are to the left. How the bristly pricker device helps capture victims in not clear. It may trigger the door and/or attract/entangle tiny creatures? “Hairs” on the stolon circled in red.
And one last oddity, number 7: Sometimes obsolete (and current) names give clues about species biology. The present name subulata, is based on the adjective “sublate,” meaning awl-shaped in reference to those inadequate-looking leaves. The older name “cleistogama” is more revealing. Cleistogamous (kliste-OG-ah-mus) flowers remain closed-up and secretly self-pollinate. These occur in today’s species either mixed with normal flowers or exclusively. A form with exclusively cleistogamous flowers was once named as a separate species, Utricularia cleistogama.


















































{kind=link}
{kind=link}
{kind=link}